
Dr. Stephan Getzin
Contact
Department of Ecological Modelling
Permoserstr. 15
04318 Leipzig
Germany

Research Interests
- Remotely Sensed Pattern Analysis
- Spatial Statistics
- Application of Unmanned Aerial Vehicles (UAV)
- Forest Ecology
- Biodiversity Research
- Nature Conservation
- Dryland Research
- Self-Organization of Vegetation
Research in the Media
 |
Unmanned Aerial Vehicles (UAV)
Unmanned aerial vehicles (UAV) or drones are increasingly used for mapping natural habitats. We have started working with drones in 2008,
at a time when the technology was not yet fully reliable. The forest project was covered in several media and had a
press release
at the University of Göttingen.
|
Publications
 | Getzin, S., Holch, S., Yizhaq, H. & Wiegand, K. (2022) Plant water stress, not termite herbivory, causes Namibia’s fairy circles. Perspectives in Plant Ecology, Evolution and Systematics (Link) | |
 | Getzin, S., Löns, C., Yizhaq, H., Erickson, T. E., Muñoz-Rojas, M., Huth, A. & Wiegand, K. (2021) High-resolution images and drone-based LiDAR reveal striking patterns of vegetation gaps in a wooded spinifex grassland of Western Australia. Landscape Ecology (Link) | |
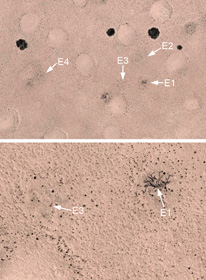 | Getzin, S., Nambwandja, A., Holch, S. & Wiegand, K. (2021) Revisiting Theron’s hypothesis on the origin of fairy circles after four decades: Euphorbias are not the cause. BMC Ecology and Evolution, 21, 102. (Link) | |
 | Getzin, S., Erickson, T. E., Yizhaq, H., Muñoz-Rojas, M., Huth, A. & Wiegand, K. (2021) Bridging ecology and physics: Australian fairy circles regenerate following model assumptions on ecohydrological feedbacks. Journal of Ecology, 109, 399-416. (Link) | |
 | Getzin, S., Yizhaq, H., Muñoz-Rojas, M., Wiegand, K. & Erickson, T. E. (2019) A multi-scale study of Australian fairy circles using soil excavations and drone-based image analysis. Ecosphere, 10(2), e02620. (Link) | |
 | Chanthorn, W., Wiegand, T., Getzin, S., Brockelman, W.Y. & Nathalang, A. (2018) Spatial patterns of local species richness reveal importance of frugivores for tropical forest diversity. Journal of Ecology, 106, 925 - 935. (Link) | |
 | Getzin, S., Fischer, R., Knapp, N. & Huth, A. (2017) Using airborne LiDAR to assess spatial heterogeneity in forest structure on Mount Kilimanjaro. Landscape Ecology, 32, 1881-1894.(Link) | |
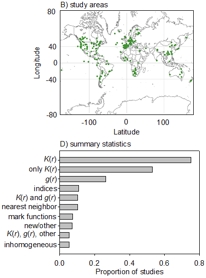 | Velázquez, E., Martínez, I., Getzin, S., Moloney, K.A. & Wiegand, T. (2016) An evaluation of the state of spatial point pattern analysis in ecology. Ecography, 39, 1042-1055. (Link) | |
 | Getzin, S., Yizhaq, H., Bell, B., Erickson, T.E., Postle, A.C., Katra, I., Tzuk, O., Zelnik, Y.R., Wiegand, K., Wiegand, T. & Meron, E. (2016) Discovery of fairy circles in Australia supports self-organization theory. PNAS, 113, 3551–3556. | |
 | Getzin, S., Wiegand, K., Wiegand, T., Yizhaq, H., von Hardenberg, J. & Meron, E. (2015) Adopting a spatially explicit perspective to study the mysterious fairy circles of Namibia. Ecography, 38, 1-11. | |
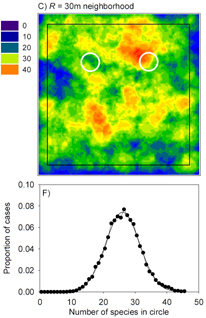 | Punchi-Manage, R., Wiegand, T., Wiegand, K., Getzin, S., Huth, A., Gunatilleke, C.V.S. & Gunatilleke, I.A.U.N. (2015) Neighborhood diversity of large trees shows independent species patterns in a mixed dipterocarp forest in Sri Lanka. Ecology, 96, 1823–1834. (Link) | |
 | Getzin, S., Nuske, R.S. & Wiegand, K. (2014) Using unmanned aerial vehicles (UAV) to quantify spatial gap patterns in forests. Remote Sensing, 6, 6988-7004. (Link) | |
 | Getzin, S., Wiegand, T. & Hubbell, S.P. (2014) Stochastically driven adult-recruit associations of tree species on Barro Colorado Island. Proceedings of the Royal Society B, 281, 20140922. (Link) | |
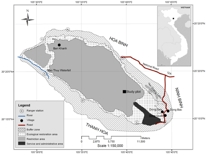 | Nguyen, H., Wiegand, K. & Getzin, S. (2014) Spatial patterns and demographics of Streblus macrophyllus trees in a tropical evergreen forest, Vietnam. Journal of Tropical Forest Science, 26, 309–319. | |
 | Nguyen, H., Wiegand, K. & Getzin, S. (2014) Spatial distributions of tropical tree species in northern Vietnam under environmentally variable site conditions. Journal of Forestry Research, 25, 257-268. | |
 | Punchi-Manage, R., Wiegand, T., Wiegand, K., Getzin, S., Gunatilleke, C.V.S. & Gunatilleke, I.A.U.N. (2014) Effect of spatial processes and topography on structuring species assemblages in a Sri Lankan dipterocarp forest. Ecology, 95, 376-386. | |
 | Zhu, Y., Getzin, S., Wiegand, T., Ren, H. & Ma, K. (2013) The relative importance of Janzen-Connell effects in influencing the spatial patterns at the Gutianshan subtropical forest. PLoS ONE, 8, e74560. | |
 | Gossner, M.M., Getzin, S., Lange, M., Pašalic, E., Türke, M., Wiegand, K., & Weisser, W.W. (2013) The importance of heterogeneity revisited from a multiscale and multitaxa approach. Biological Conservation, 166, 212-220. | |
 | Ward, D., Wiegand, K., Getzin, S. (2013) Walter's two-layer hypothesis revisited: back to the roots! Oecologia, 172, 617-630. | |
 | Punchi-Manage, R., Getzin, S., Wiegand, T., Kanagaraj, R., Gunatilleke, C.V.S., Gunatilleke, I.A.U N., Wiegand, K. & Huth, A. (2013) Effects of topography on structuring local species assemblages in a Sri Lankan mixed dipterocarp forest. Journal of Ecology, 101, 149-160. (Link)
| |
 | Wiegand, T., Huth, A., Getzin, S., Wang, X., Hao, Z., Gunatilleke, C.V.S. & Gunatilleke, I.A.U N. (2012) Testing the independent species' arrangement assertion made by theories of stochastic geometry of biodiversity. Proceedings of the Royal Society B, 279, 3312-3320. (Link)
| |
 | Getzin, S., Wiegand, K. & Schoening, I. (2012) Assessing biodiversity in forests using very high-resolution images and unmanned aerial vehicles. Methods in Ecology and Evolution, 3, 397-404. (Link)
| |
 | Lan, G., Getzin, S., Wiegand, T., Hu, Y., Xie, G., Zhu, H. & Cao, M. (2012) Spatial distribution and interspecific associations of tree species in a tropical seasonal rain forest of China. PLoS ONE, 7, e46074. | |
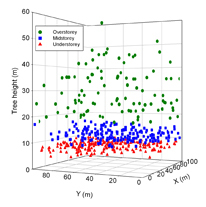 | Getzin, S. , Worbes, M., Wiegand, T. & Wiegand, K. (2011) Size dominance regulates tree spacing more than competition within height classes in tropical Cameroon. Journal of Tropical Ecology, 27, 93-102. | |
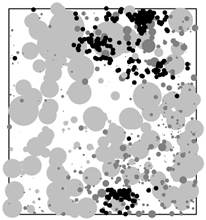 | Getzin, S. , Wiegand, T., Wiegand, K. & He, F. (2008) Heterogeneity influences spatial patterns and demographics in forest stands. Journal of Ecology, 96, 807-820. (Link)
| |
 | Moustakas, A., Wiegand, K., Getzin, S. , Ward, D., Meyer, K.M., Guenther, M. & Mueller, K.-H. (2008) Spacing patterns of an Acacia tree in the Kalahari over a 61-year period: how clumped becomes regular and vice versa. Acta Oecologica, 33, 355-364. | |
 | Getzin, S. , Wiegand, K., Schumacher, J. & Gougeon, F.A. (2008) Scale-dependent competition at the stand level assessed from crown areas. Forest Ecology and Management, 255, 2478-2485. | |
 | Getzin, S. & Wiegand, K. (2007) Asymmetric tree growth at the stand level: Random crown patterns and the response to slope. Forest Ecology and Management, 242, 165-174. | |
 | Getzin, S. (2007) Structural Fire Effects in the World's Savannas. A Synthesis for Biodiversity and Land-Use Managers. VDM Verlag, Saarbruecken. Book-ISBN: 978-3-8364-3664-9 | |
 | Getzin, S. , Dean, C., He, F., Trofymow, J.A., Wiegand, K. & Wiegand, T. (2006) Spatial patterns and competition of tree species in a Douglas-fir chronosequence on Vancouver Island. Ecography, 29, 671-682. (Link)
| |
 | Getzin, S. (2005) The suitability of the degradation gradient method in arid Namibia. African Journal of Ecology, 43, 340-351. | |
 | Becker, T. & Getzin, S. (2000) The fairy circles of Kaokoland (North-West-Namibia) - origin, distribution, and characteristics. Basic and Applied Ecology, 1, 149-159. |